
L18-MDP
-
Cat.code:
tlrl-lmdp
- Documents
ABOUT
NOD2 Agonist - Muramyl dipeptide with a C18 fatty acid chain
L18-MDP is a potent agonist of the cytosolic receptor NOD2. It is a lipophilic derivative of muramyl dipeptide (MDP) in which the OH group of the C6 position is esterified with stearic acid. MDP is the minimal bioactive peptidoglycan motif present in almost all bacteria. NOD2 acts as a general sensor of bacterial invasion through MDP detection [1].
Mode of action:
In an attempt to enhance the protective activity of MDP, numerous derivatives were synthesized. Among them, L18-MDP, a 6-O-acyl derivative with a stearoyl fatty acid, showed the highest activity [2]. This modification results in enhanced uptake of MDP and hydrolysis of the ester bond within the cell, releasing MDP in the cytoplasm. NOD2 activation with L18-MDP causes pro-inflammatory cytokine release through the mitogen-activated protein kinase (MAPK) and NF-κB activation, thus contributing to host defense [3].
In HEK-Blue™ NOD2 cells, L18-MDP is more potent than MDP at inducing human NOD2 responses (see figures).
Key features:
- Potent NOD2 agonist
- Synthetic lipophilic derivative of MDP
- Each lot is functionally tested
![]() Read our review on NOD-Like Receptors
Read our review on NOD-Like Receptors
References:
1. Girardin SE. et al., 2003. Nod2 is a general sensor of peptidoglycan through muramyl dipeptide (MDP) detection. J Biol Chem. 278(11):8869-72.
2. Matsumoto K. et al., 1981. Stimulation of nonspecific resistance to infection induced by 6-O-acyl muramyl dipeptide analogs in mice. Infect Immun. 32(2):748-58.
3. Fujimura T. et al., 2011. A synthetic NOD2 agonist, muramyl dipeptide (MDP)-Lys (L18) and IFN-β synergistically induce dendritic cell maturation with augmented IL-12 production and suppress melanoma growth. J Dermatol Sci. 62(2):107-15.
All products are for internal research use only, and not for human or veterinary use.

SPECIFICATIONS
Specifications
C37H66N4O12
1-100 ng/ml
1 mg/ml in water
0.2 µm filtration
NOD2 activation in cellular assays
Activation of NOD2 confirmed using HEK-Blue™ NOD2 cells, Absence of NOD1 activity confirmed using HEK‑Blue™ NOD1 cells, Absence of bacterial contamination confirmed using HEK-Blue™ TLR2 and HEK‑Blue™ TLR4 cells
CONTENTS
Contents
-
Product:L18-MDP
-
Cat code:tlrl-lmdp
-
Quantity:1 mg
1.5 ml sterile endotoxin-free water
Shipping & Storage
- Shipping method: Room temperature
- -20°C
- Avoid repeated freeze-thaw cycles
Storage:
Caution:
Details
NOD1 and NOD2
The cytosolic NOD-Like Receptors (NLRs, also known as NODs or NALP) are Nucleotide-binding Oligomerization Domain containing receptors. To date, 22 NLRs have been identified in humans and constitute a major class of intracellular pattern recognition receptors (PRRs) [1].
The founding NLR-family members NOD1 (CARD4) and NOD2 (CARD15) recognize distinct motifs of peptidoglycan (PGN), an essential constituent of the bacterial cell wall. NOD1 senses the D-γ-glutamyl-meso-DAP dipeptide (iE-DAP), which is found in PGN of all Gram-negative and certain Gram-positive bacteria [1, 2] whereas NOD2 recognizes the muramyl dipeptide (MDP) structure found in almost all bacteria. Thus NOD2 acts as a general sensor of PGN and NOD1 is involved in the recognition of a specific subset of bacteria. Both iE-DAP and MDP must be delivered intracellularly either by bacteria that invade the cell or through other cellular uptake mechanisms. Ligand-bound NOD1 and NOD2 oligomerize and signal via the serine/threonine RIP2 kinase through CARD-CARD homophilic interactions [3]. Once activated, RIP2 mediates ubiquitination of NEMO/IKKγ leading to the activation of NF-κB and the production of inflammatory cytokines. Furthermore, poly-ubiquitinated RIP2 recruits TAK1, which leads to IKK complex activation and the activation of MAPKs [4].
Genetic mutations in NOD2 are associated with Crohn’s disease, a chronic inflammatory bowel disease [5]. In addition, numerous studies have recently revealed that NOD1 and NOD2 have a close relationship with a variety of cancers via controlling proliferation, altering immunosurveillance, and interacting with tissue bacteria, including intestinal commensal intestinal microflora. Moreover, additional research into the mechanisms of NOD1 and NOD2 in cancers would shed light on the innate immunity-cancer relationship and provide intriguing targets for immunotherapy [6].
References:
1. Chamaillard M. et al., 2003. An essential role for NOD1 in host recognition of bacterial peptidoglycan containing diaminopimelic acid. Nat. Immunol. 4: 702-707.
2. Girardin S. et al., 2003. Nod1 detects a unique muropeptide from Gram-negative bacterial peptidoglycan. Science 300: 1584-1587.
3. Kobayash, K. et al., 2002. RICK/Rip2/CARDIAK mediates signalling for receptors of the innate and adaptive immune systems. Nature 416: 194-199.
4. Kobayashi K. et al., 2005. Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science 307: 731-734.
5. Ogura Y. et al., 2001. A frameshift mutation in NOD2 associated with susceptibility to Crohn’s disease. Nature 411: 603-606.
6. Wang D., 2022. NOD1 and NOD2 Are Potential Therapeutic Targets for Cancer Immunotherapy. Comput Intell Neurosci.;2022:2271788.
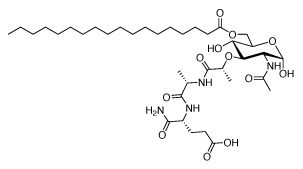
Structure of L18-MDP:
DOCUMENTS
Documents
Technical Data Sheet
Safety Data Sheet
Validation Data Sheet
Certificate of analysis
Need a CoA ?
 Powered by Bioz
Powered by Bioz